
Сокращения и аббревиатуры
Bmp, костный морфогенетический белок; 2,3 - DPG, 2,3 - дифосфоглицерат;
E#, день эмбрионального развития, где число указывает возраст эмбриона в днях после зачатия;
EPO, эритропоэтин;
fL, фемтолитр;
Gata1, 2 и 4, GATA - связывающие белки 1, 2 и 4;
HSC, гемопоэтические стволовые клетки;
Ihh, индийский еж;
IL, интерлейкин;
P#, день постнатального развития, где число указывает возраст новорожденного в днях после родов; pg, пикограмм;
PU.1, пуриновый бокс - связывающий фактор транскрипции;
Scl/Tal - 1, лейкоз стволовых клеток/T - фактор острого лейкоза.
ОСНОВНЫЕ ПРИНЦИПЫ РАЗВИТИЯ ГЕМОПОЭТИЧЕСКОЙ СИСТЕМЫ
Клетки крови, вырабатываемые на разных стадиях развития, различаются по морфологии и функциям. Таким образом, примитивные («фетальные») клетки, вырабатываемые на ранних стадиях гестации, имеют заметно отличающиеся свойства от своих окончательных («взрослых») аналогов, вырабатываемых на поздних стадиях гестации и в постнатальной жизни. Этот принцип наиболее полно охарактеризован в клетках эритроидной линии. Примитивные эритроциты (эритроциты) образуются в желточном мешке, тогда как окончательные эритроциты вырабатываются печенью, а затем селезенкой и костным мозгом. Примитивные эритроциты образуют ядра в кровотоке примерно до 12,5 дня (E12.5) беременности, после чего ядра постепенно конденсируются, прежде чем будут сброшены между E14.5 и E16.5. Энуклеированные примитивные эритроциты сохраняют свой большой размер и могут оставаться в кровотоке вплоть до 5-го дня после рождения (P5). Как примитивные, так и дефинитивные эритроциты высвобождаются в течение большей части второй половины беременности (с E10 по E18), хотя соотношение меняется с течением времени от преимущественно примитивных к преимущественно дефинитивным эритроцитам.
Примитивные и дефинитивные эритроциты можно различить по их размеру. Объем примитивных эритроцитов варьируется от 465 до 530 фемтолитров (fL), что примерно в шесть раз больше, чем у дефинитивных эритроцитов. Содержание гемоглобина в примитивных эритроцитах, от 80 до 100 пикограмм (пг)/клетку, также почти в шесть раз превышает количество, обнаруженное в дефинитивных эритроцитах. Как примитивные, так и дефинитивные эритроциты имеют базофильную цитоплазму, когда они впервые производятся из-за обильного шероховатого эндоплазматического ретикулума, но базофилия отступает по мере достижения максимального содержания гемоглобина.
ПРИМИТИВНЫЙ ГЕМОПОЭЗ
Эритроидные клетки
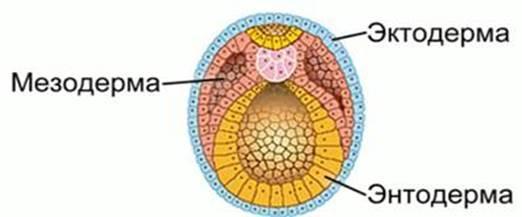
Гемопоэз происходит в нескольких местах внутри эмбриона и во внеэмбриональных тканях. Первая фаза производства клеток крови, называемая примитивным гемопоэзом, отвечает за выработку элементов крови на самой ранней стадии эмбриогенеза. Примитивный гемопоэз происходит в висцеральном желточном мешке, начиная примерно с E7.0. Таким образом, примитивные гемопоэтические клетки являются одними из самых ранних отдельных тканей, которые дифференцируются в эмбрионе. Образование примитивных клеток быстро снижается после E11. Висцеральный желточный мешок или внеэмбриональная спланхноплевра (термин для структуры, в которой мезодерма и энтодерма непосредственно противопоставлены) возникает в результате миграции внеэмбриональной мезодермы, текущей из каудальной первичной полоски вдоль внутренней поверхности висцеральной энтодермы. Мезодермальные клетки, призванные инициировать и поддерживать кроветворение, были названы гемангиобластами, поскольку смежность примитивного кроветворения и васкулогенеза как в пространстве, так и во времени предполагает, что примитивные кроветворные и эндотелиальные клетки в желточном мешке имеют общего предка. Гемангиобласты возникают как недифференцированные клетки на стадии примитивной полоски и обязуются производить определенную клеточную линию до образования кровяных островков. Эти плюрипотентные клетки также могут дифференцироваться в другие ткани, происходящие из мезенхимы.
Между E7.5 и E9.0 гемангиобласты образуют множественные агрегаты, называемые кровяными островками. Каждый кровяной островок содержит центральное ядро из неприкрепленных внутренних гемангиобластов (кроветворных предшественников), окруженное ободом из веретенообразных внешних гемангиобластов (эндотелиальных предшественников). Ядросодержащие эритроидные клетки впервые распознаются в ядрах кровяных островков на E8.0 и становятся очевидными в циркуляции в сердечно-сосудистой системе, начиная с E8.25. На этой стадии эмбриональные эритробласты попадают в кровообращение, где они продолжают делиться примерно до E13.0.
Большинство клеток, образующихся во время примитивного кроветворения, относятся к эритроидной линии. Приверженные эритроидные колониеобразующие клетки поступают в желточный мешок примерно на E7.25. Эти клетки расширяются до E8.0, а затем дифференцируются в примитивные эритробласты; все колониеобразующие клетки полностью регрессируют к E9.0,34 что примерно соответствует самой ранней фазе дефинитивного эритропоэза. Примитивные эритробласты служат единственным источником эритроцитов у раннего эмбриона с E8.0 до примерно E10.534 и остаются важным источником эритроцитов до E13. Таким образом, эмбрионы с возрастом развития между E8.0 и E11, страдающие анемией, страдают дефектом примитивного эритропоэза. Интересно, что кажущиеся глубокими дефекты примитивного кроветворения, приводящие к стойким функциональным нарушениям во взрослом возрасте, могут и не вызывать аномального гематологического профиля у эмбриона.
Другие клетки
Недавние исследования показывают, что другие линии гемопоэтических клеток также генерируются в желточном мешке во время этой примитивной стадии развития кроветворения. Примитивные лимфоидные предшественники и даже некоторые взрослые стволовые клетки развиваются на E7.5 и впоследствии засеивают другие места кроветворения, включая область аорты - гонады - мезонефроса (AGM), пупочные сосуды и печень. Примитивные макрофаги были идентифицированы в желточном мешке на E8.0 4 - E9.0. Эксперименты in vitro продемонстрировали, что клетки желточного мешка E7.5 могут давать начало функциональным мегакариоцитарным предшественникам на E10.5. Многие гемангиобласты фактически служат би- или олиго-потентными предшественниками, включая те, которые способны к приверженности эритроцитарно-миелоидным, эритроцитарно-мегакариоцитарным, гранулоцитарно-макрофагальным и лимфоидным (В-клетки и Т-клетки)/миелоидным линиям. Также сообщалось, что стволовые клетки для тучных клеток возникают в желточном мешке во время примитивного кроветворения.
ДЕФИНИТИВНЫЙ ГЕМОПОЭЗ
Независимо от их исходного места синтеза de novo, HSC мигрируют в другие места, которые поддерживают дефинитивный гемопоэз: эмбриональная печень, затем эмбриональный тимус, фетальная селезенка и костный мозг (в этом порядке). Эти последние места назначения не производят HSC de novo, а скорее содержат ниши, подходящие для расширения вновь прибывших HSC. Пригодность таких ниш контролируется специфическими характеристиками их стромальных поддерживающих клеток. Эмбриональная печень колонизируется первой, по-видимому, потому, что она разделяет многие молекулярные и функциональные сходства с желточным мешком. Она обеспечивает основной локус для дефинитивного гемопоэза с E12 по E16. HSC попадают в эмбриональную печень несколькими последовательными волнами между E9.0 или E10.0 и E12. Первые HSC, попавшие в печень, являются плюрипотентными и могут образовывать любой тип гемопоэтических клеток. Их первым шагом во внутрипеченочном созревании является приверженность более ограниченному диапазону вариантов линий, как правило, как эритромиелоидный предшественник или общий миелолимфоидный предшественник. Окончательные эритроидные предшественники созревают и энуклеируются в эритроидных островках в печени перед тем, как попасть в кровоток. Миелолимфоидные предшественники, полученные из печени, впоследствии развиваются в бипотентные клетки (В-клетки и миелоидные или Т-клетки и миелоидные), прежде чем приступить к производству одной клеточной линии. Некоторые предшественники Т-клеток имеют бипотентную приверженность к линии NK-клеток. Предшественники Т-клеток, предназначенные для переноса в эмбриональный тимус, вырабатываются даже у бестимусных мышей, что указывает на то, что фетальная печень может играть роль в содействии ранней дифференцировке Т-клеток.
Эмбриональный тимус и фетальная селезенка засеваются либо из печени, либо из AGM, либо из обоих, начиная примерно с E13 для тимуса и с E15 для селезенки. Тимус обычно принимает только те HSC, которые предназначены для производства Т-клеток, тогда как другие мультипотентные миелолимфоидные элементы направляются в другие места. Количество предшественников Т-клеток в печени обильно на E12, но затем уменьшается, тогда как популяция внутрипеченочных предшественников В-клеток демонстрирует обратную тенденцию. Большинство типов дефинитивных гемопоэтических клеток в селезенке возникают из клеток-предшественников, которые предназначены для определенной линии перед тем, как покинуть печень. Мультипотентные HSC, попадающие в селезенку, прекращают пролиферацию и дифференцируются в зрелые макрофаги. Эти клетки могут регулировать внутриселезеночный эритропоэз.
Костный мозг впервые получает HSC из печеночных депо примерно на E16. После этого распределение колониеобразующих гемопоэтических предшественников смещается от преимущественно гепатоцентрической локализации на E18 через более дисперсное распределение (костный мозг, печень и селезенка примерно в равных количествах) на P2 к профилю, благоприятствующему костному мозгу и в меньшей степени селезенке на P4 и позже. Таким образом, костный мозг, печень и селезенка функционируют совместно, регулируя окончательный гемопоэз. Во время сотрудничества каждый орган поддерживает молекулярно отличную подгруппу гемопоэтических предшественников.
Активные гемопоэтические клетки-предшественники необходимы для развития всех типов клеток крови, наблюдаемых у взрослых животных. Эти клетки образуются в ходе дефинитивного кроветворения, которое представляет собой процесс образования клеток крови в костном мозге. Гемопоэтические стволовые клетки (HSC), полученные из области AGM, участвуют во всех основных линиях клеток крови, включая эритроидные (красные кровяные тельца), лимфоидные (В- и Т-клетки) и миелоидные (гранулоциты, моноциты, макрофаги) клетки. Популяция HSC, полученная из плаценты, поддерживает выработку эритроцитов, В- и Т-лимфоцитов и миелоидных клеток. В отличие от этого, HSC, полученные из желточного мешка, способствуют образованию только лимфоидных и миелоидных типов клеток. Еще предстоит определить, проявляют ли предшественники определенных линий клеток крови различные функциональные и молекулярные характеристики на поздних стадиях внутриутробной и послеродовой жизни.
Эмбрионы поздней стадии (E13–E15), плоды (E16 до рождения) и новорожденные, у которых проявляется анемия, страдают дефектом дефинитивного эритропоэза. Аномалии, связанные с этим проявлением, включают полное отсутствие дефинитивного кроветворения и неспособность клеток-предшественников должным образом колонизировать внутриэмбриональные участки кроветворения. Могут быть затронуты множественные клеточные линии; такой комбинированный эффект предполагает, что гемопоэтический дефект возникает в би- или мультипотентной стволовой клетке, а не в той, которая предназначена для формирования определенной клеточной линии. Проявление анемии поздней стадии также может быть результатом общей задержки роста и развития, а не целенаправленной аномалии в эритроцитарной линии.
У молодых животных количество циркулирующих кровяных клеток отличается от количества взрослых. Количество эритроцитов увеличивается более чем вдвое между рождением и ранней взрослой жизнью. Количество циркулирующих лейкоцитов при рождении составляет приблизительно 20% от уровня взрослых, а затем увеличивается до взрослого уровня к 6–7 неделям. Количество тромбоцитов у новорожденных составляет приблизительно одну треть.
ТРАНСФОРМАЦИЯ ГЕМОГЛОБИНА
Степень экспрессии отдельных генов глобина и сочетание экспрессируемых генов глобина со временем меняются. Например, энуклеированные примитивные эритроциты содержат относительно больше β min, чем дефинитивные эритроциты. На E11.5 β min составляет приблизительно 80% β-глобина в кровотоке. Этот уровень снижается приблизительно на 60% при рождении. Примитивные эритроциты экспрессируют увеличивающиеся уровни взрослых глобинов по мере развития беременности, тогда как дефинитивные эритроциты содержат только варианты взрослых белков. Эта эволюция указывает на то, что характер экспрессии глобина переключается по мере замены примитивных эритроцитов дефинитивными эритроцитами. Молекулярные механизмы, которые регулируют процесс переключения, сложны. Время этого переключения между E10.5 и E12.5 совпадает с первоначальным подъемом дефинитивного эритропоэза. Нарушение синхронизации этого переключения характерно для некоторых моделей заболеваний кроветворной системы на мышах.
Успешное поддержание развивающегося зародыша зависит от преимущественного захвата кислорода в эмбриональных и фетальных тканях. Поэтому примитивные эритроциты, как правило, имеют более высокое сродство к кислороду, чем материнские эритроциты, хотя домашние кошки являются исключением. Эта секвестрация кислорода опосредована двумя основными механизмами. Механизм, относящийся к раннему эмбриональному периоду, заключается в большем сродстве эмбрионального гемоглобина в примитивных эритроцитах к кислороду по сравнению с гемоглобином взрослого организма. С другой стороны, дефинитивные эритроциты в позднем эмбрионе и плоде обладают более низкой концентрацией 2,3-дифосфоглицерата (2,3-DPG), чем материнские эритроциты. Более высокие уровни 2,3-DPG облегчают высвобождение кислорода в ткани. После рождения уровни содержания 2,3-DPG в эритроцитах повышаются до уровней взрослых в течение 10-15 дней.
МОЛЕКУЛЯРНЫЕ МЕХАНИЗМЫ, РЕГУЛИРУЮЩИЕ РАЗВИТИЕ КРОВЕТВОРЕНИЯ
Секретируемые молекулы также являются важными регуляторами развития кроветворения во время беременности. Например, эритропоэтин (EPO) поддерживает как примитивный, так и дефинитивный эритропоэз, стимулируя пролиферацию и дифференциацию незрелых примитивных и дефинитивных эритроцитов. Снижение активности EPO в желточном мешке значительно снижает количество колониеобразующих клеток и эритробластов посредством чрезмерного апоптоза. Тромбопоэтин выполняет аналогичную функцию для мегакариоцитов, хотя также требуются другие цитокины (интерлейкин-3 [ИЛ-3], ИЛ-6) и факторы роста (гранулоцитарный колониестимулирующий фактор, фактор стволовых клеток). Другие сигнальные пути лиганда/рецептора, которые, как показано, влияют на развитие кроветворения, включают молекулу индийского ежа (molecule Indian hedgehog) (Ihh), полученную из эндодермы, и костный морфогенетический белок 4 (Bmp4), оба из которых участвуют в образовании кровяных островков и васкулогенезе в желточном мешке. В целом, секретируемые молекулы действуют посредством взаимодействия со специфическим фактором транскрипции
Молекулы клеточной адгезии семейства интегринов необходимы для правильной миграции гемопоэтических предшественников. Например, β1 - интегрины необходимы для того, чтобы HSC достигали печени эмбриона, а затем селезенки и костного мозга плода на соответствующих стадиях развития.Потеря β1 - интегринов предотвращает адгезивное взаимодействие между HSCs и эндотелиальными клетками, тем самым блокируя HSCs в сосудах.Некоторые интегрины выполняют дополнительные функции, помимо своей целевой функции. Потеря β1 - интегринов предотвращает адгезивное взаимодействие между HSCs и эндотелиальными клетками, тем самым блокируя HSCs в сосудах.Некоторые интегрины выполняют дополнительные функции, помимо своей целевой функции. Например, β4 - интегрины необходимы не только для правильного самонаведения, но и для размножения и дифференцировки предшественников эритроидных и В-клеток в печени, селезенке и костном мозге. Как и в случае с секретируемыми факторами, активность некоторых интегринов в большей степени относится к поздним срокам беременности и неонатальным стадиям, а не к более ранним стадиям развития кроветворения. Эта хронология была задокументирована для β4 - интегрина в отношении лимфоидной и миелоидной дифференцировки.
Сокращения и аббревиатуры
RBC — эритроцит;
WBC — белые кровяные клетки.
функциональные взаимоотношения между различными компонентами костного мозга. Более подробное обсуждение биохимического и молекулярного контроля кроветворения и кроветворной микросреды читатель может найти в главах 1 и 5.
ПОДДЕРЖИВАЮЩИЕ СТРУКТУРЫ
Слой эндостальных клеток толщиной от одной до двух клеток с тонким слоем соединительной ткани выстилает все костные поверхности внутри костномозговой полости. Этот слой иногда перемежается остеобластами и остеокластами костной ткани и может быть пересечен эндостальными кровеносными сосудами, соединяющими кроветворное пространство с костью. Остеобласты участвуют в образовании костной ткани и происходят из мультипотентных мезенхимальных стромальных клеток-предшественников, которые также дают начало стромальным клеткам костного мозга и адипоцитам. Остеокласты - это многоядерные клетки, происходящие из слившихся моноцитарно- макрофагальных предшественников под влиянием многочисленных сигналов, в том числе исходящих от остеобластов. Остеобласты и остеокласты реконструируют кость в пространстве костного мозга, влияя на эндостальную среду и, вероятно, способствуя регуляции пролиферации гемопоэтических стволовых клеток и их транспортировке. Остеобласты и остеокласты также продуцируют кроветворные цитокины, и взаимодействие между костью и кроветворными клетками может влиять на обновление и ремоделирование костной ткани.
Тонкие, веретеновидные или звездчатые стромальные клетки распространяются от эндостальных областей в паренхиму кроветворной ткани. Эти клетки, вероятно, происходят из фибробластов костного мозга и образуют поддерживающую сеть для кроветворных клеток, жировой ткани и кровеносных сосудов. Стромальные клетки вырабатывают растворимые факторы, которые способствуют регуляции кроветворения, и взаимодействуют с кроветворными клетками-предшественниками посредством прямого межклеточного контакта. Стромальные клетки костного мозга вырабатывают структурные фибриллы, такие как коллаген, ретикулин, ламинин и фибронектин, а также основное вещество, состоящее из воды, солей, гликозаминогликанов и гликопротеинов, которые в совокупности называются внеклеточным матриксом. Как и поддерживающие клеточные структуры костного мозга, внеклеточный матрикс участвует как в структурной, так и в биохимической поддержке кроветворения.
Костный мозг содержит преимущественно коллаген I и III типов, которые принимают свою окончательную форму после секреции во внеклеточное пространство, где они подвергаются ферментативной модификации. Ретикулин отличается от коллагена в костном мозге наличием тонких аргирофильных волокон, которые состоят из ядра коллагена I типа, окруженного волокнами коллагена III типа, встроенными в матрицу гликопротеинов и гликозаминогликанов. Хотя тонкая соединительнотканная структура костного мозга не видна на обычно обрабатываемых гистологических срезах, специальные окраски могут улучшить ее визуализацию. Коллаген можно визуализировать с помощью трихромного окрашивания по Маллори или Массону, тогда как серебряное окрашивание по Гомори выявляет наличие ретикулина. Это может быть важно при патологических состояниях, при которых избыточная выработка матриксного материала способствует возникновению заболевания. Дифференциация коллагенового и ретикулинового фиброза может иметь диагностическое значение и влиять на вероятность обратимости поражения. Расширенный внеклеточный матрикс характерен и для других заболеваний костного мозга, выходящих за рамки классических синдромов миелофиброза.
Жировая ткань, перемежающаяся с кроветворной тканью, опутана одними и теми же опорными структурами. Связь между образованием костной и жировой ткани до конца не изучена. Адипоциты, вероятно, происходят из тех же мезенхимальных предшественников, которые производят стромальные клетки и остеобласты, и имеются некоторые доказательства взаимопревращения клеток, происходящих из комитированных остеобластических и адипогенных линий, полученных из популяции мезенхимальных предшественников. Адипоциты — самые многочисленные стромальные клетки костного мозга. У здоровых животных жировая ткань занимает примерно от 25% до 75% объема костного мозга в зависимости от возраста животного. Жировая ткань также содержит
другие типы клеток, которые менее заметны, чем адипоциты, на обычных гистологических срезах. Эти клетки также вносят вклад в структурную и функциональную роль жировой ткани в кроветворении и включают эндотелиальные клетки, макрофаги и клетки-предшественники адипоцитов. В костном мозге присутствует как бурая, так и белая жировая ткань; Различия в биологической функции этих типов жиров до конца не изучены. Жировая ткань костного мозга, как правило, относительно устойчива к липолизу во время голодания по сравнению с жировой тканью в других частях тела.
Помимо обеспечения структурной поддержки, жировая ткань также может участвовать в кроветворной микросреде. Клетки, полученные из жировой ткани костного мозга, способны поддерживать дифференцировку кроветворных предшественников in vitro. Важны также эндокринные и паракринные функции жировой ткани. Адипокины — биологически активные вещества, вырабатываемые жировой тканью, включают регуляторы кроветворения и иммунного ответа.
СОСУДИСТАЯ СЕТЬ И СТРОЕНИЕ СИНУСОВ

обзор. Exp Hematol 1981;9:391 – 410, с
разрешение.)
Кроветворение происходит во внесосудистых пространствах между венозными синусами и имеет тесную морфологическую и функциональную взаимосвязь с клетками, выстилающими венозные синусы. Венозные синусы выстланы полным просветным слоем широких плоских эндотелиальных клеток и неполным наружным слоем аблуминальных ретикулярных клеток (рис. 2.2). Ретикулярные клетки поддерживают тесные физические связи с гемопоэтическими клетками, расположенными вблизи стенок синуса, часто обволакивая или иным образом контактируя с гемопоэтическими предшественниками. Базальная пластинка между синусоидами и кроветворным пространством тонкая и прерывистая, что облегчает попадание зрелых кроветворных клеток в кровоток. Эндотелиальные клетки синуса могут регулировать перемещение клеток и других веществ в системный кровоток. Ультраструктурно эндотелиальные клетки синуса имеют четкие клеточные соединения, которые не являются плотными, и наблюдался выход гемопоэтических клеток через миграционные поры в эндотелиальных клетках. Другие твердые частицы могут проникать в эндотелиальные клетки синуса в процессе эндоцитоза.
ИННЕРВАЦИЯ
Синусоидальная система менее богата иннервацией, чем артериальная сосудистая сеть, при этом нервные окончания часто контактируют со стенками синусоидов. Другие нервные волокна, по-видимому, заканчиваются в пределах кроветворной материнской клетки.
химусе или вдоль эндоста. Неясно, существует ли общая или раздельная иннервация костной и кроветворной ткани, хотя по крайней мере некоторые нервные волокна в костном мозге, по-видимому, происходят из минерализованной кости. Костный мозг содержит эфферентные норадренергические и пептидергические симпатические и предполагаемые чувствительные нервные волокна. Имеются некоторые данные о том, что сигналы симпатической нервной системы могут способствовать регуляции кроветворения, иммунной функции и миграции гемопоэтических стволовых клеток.

КЛЕТОЧНАЯ ОРГАНИЗАЦИЯ
Гемопоэтические клетки происходят из общей плюрипотентной стволовой клетки, которая дает начало лимфоидным и миелоидным клеткам-предшественникам. Лимфоидные клетки-предшественники генерируют потомство лимфоцитов, тогда как миелоидные клетки-предшественники генерируют эритроидные клетки, мегакариоциты, базофилы, эозинофилы и общую гранулоцитарно-макрофагальную клетку, которая продуцирует нейтрофилы и макрофаги. Каждая клеточная линия демонстрирует пирамидальную прогрессию числа клеток, при этом наименее зрелые клетки представлены в наименьшем количестве, а клетки на последующих стадиях развития представлены в возрастающих пропорциях.
Мегакариоциты
Развитие мегакариоцитов начинается с мегакариобласта, прогрессирует до промегакариоцитов и базофильных мегакариоцитов и завершается образованием зрелых мегакариоцитов. Предшественники мегакариоцитов постепенно увеличиваются по мере дифференциации, становясь самой крупной клеткой в костном мозге. Их ядра созревают из одного ядра в большое многодольчатое ядро посредством процесса, называемого эндомитозом, который представляет собой репликацию ДНК без клеточного деления. Цитоплазма ранних предшественников мегакариоцитов сканируется и становится глубоко базофильной, становясь более обильной, слегка базофильной и заполненной многочисленными эозинофильными гранулами по мере созревания клетки.
По мере созревания мегакариоциты мигрируют к венозным синусам и могут составлять часть слоя эндотелиальных клеток. Такое расположение позволяет цитоплазматическим отросткам распространяться через эндотелиальные щели и выбрасывать протромбоциты непосредственно в просвет синуса. Тромбоциты высвобождаются из протромбоцитарных отростков в периферическое кровообращение (см. Главу 9).
Эритробластические (рубробластические) островки
Стадии эритропоэза включают рубробласты, прорубрициты, рубрициты, метарубрициты, ретикулоциты и зрелые эритроциты (см. Главу 6). По мере созревания эритроидных предшественников клетки становятся меньше, ядерно-цитоплазматическое соотношение уменьшается, цитоплазма становится менее базофильной и более полихроматофильной, а ядерный хроматин уплотняется. У млекопитающих ядро выдавливается перед созреванием в зрелую клетку. Эритропоэз происходит в отдельных эритробластических островках (рис. 2.3), которые представляют собой скопления клеток, которые иногда можно наблюдать в цитологических образцах костного мозга. Эритробластические островки образуются вокруг центрального макрофага, который проецирует мембранные отростки для содействия эритропоэзу, обеспечивая железом и, вероятно, другими питательными веществами и гемопоэтическими цитокинами. Эти макрофаги также фагоцитируют вытесненные ядра и дефектные клетки. Эритроидное потомство находится в концентрических кругах, окружающих центральный макрофаг, а более молодые формы находятся ближе к макрофагу. Центральные макрофаги набираются из подгруппы резидентных макрофагов, полученных из предшественников моноцитов. Эритробластические островки расположены вблизи венозных синусов. Недавнее исследование эритропоэза в костном мозге крыс предполагает, что эритробластические островки подвижны и мигрируют к синусоидам по мере созревания, и что эритробластические островки состоят из клеток на схожих стадиях развития эритроидов.

Компартментализация или компартментация - разделение клеток эукариот на отсеки (компартменты), покрытые оболочкой из бислоя липидов (т.е. отделенные мембраной), в которых локализованы определенные биохимические процессы. Примерами компартментов могут быть митохондрии, хлоропласты, пероксисомы, лизосомы, эндоплазматический ретикулум, ядро клетки и аппарат Гольджи.
Сокращения и аббревиатуры
ОПРЕДЕЛЕНИЕ СТВОЛОВЫХ КЛЕТОК
Характеристики
Стволовые клетки представляют собой популяцию неспециализированных клеток-предшественников, которые обладают способностью к самообновлению и способностью к дифференцировке, что приводит к образованию зрелых клеток и тканей. Последняя функция отчетливо проявляется в эмбриональных стволовых клетках (EPC), поскольку они приводят к образованию многочисленных различных клеток и тканей зрелого организма. Небольшое количество стволовых клеток сохраняется на протяжении всей жизни в виде взрослых стволовых клеток и является резервуаром для замены короткоживущих клеток или регенерации поврежденных тканей. Гемопоэтические стволовые клетки (HSC) являются резервуаром для замены клеток крови и присутствуют с частотой 1 на каждые 10 000–100 000 клеток крови. (рис. 3.1; см. главы 6–10).
При определении стволовых клеток используются две общие функциональные характеристики. Первая из них — способность к долгосрочному самообновлению. Стволовые клетки обладают способностью посредством митотического деления поддерживать популяцию недифференцированных клеток в пуле стволовых клеток в течение месяцев и лет на протяжении многих циклов клеточного деления. При делении стволовых клеток в среднем одна дочерняя клетка является репликой и остается в недифференцированном состоянии, тогда как вторая дочерняя клетка запрограммирована на дифференциацию. Такое образование двух дочерних клеток с разными свойствами называется асимметричным делением клетки. Вторая характеристика стволовых клеток — способность образовывать дифференцированные или специализированные типы клеток.
Потентность — это термин, который используется для описания степени или масштаба, в котором могут быть сформированы множественные функциональные клеточные линии. Тотипотентные стволовые клетки — это клетки, способные формировать целые организмы, включая дополнительные эмбриональные ткани (например, плаценту). Этот тип стволовых клеток может быть получен из зиготы или раннего бластомера. Плюрипотентные стволовые клетки способны образовывать все типы клеток организма (например, эмбриональные стволовые клетки). Мультипотентные стволовые клетки генерируют все дифференцированные клетки определенной линии (например, ГСК) и будут представлять особый интерес для тем этого текста. Наконец, унипотентные стволовые клетки дают начало только одной клеточной линии (например, сперматогониальным стволовым клеткам).

Тесты и маркеры
Функциональные анализы, демонстрирующие плюрипотентную или мультипотентную природу стволовых клеток, были проведены с помощью in vitro формирования эмбриональных тел, in vivo генерации тератом у мышей с тяжелым комбинированным иммунодефицитом (SCID, severe combined immunodefi ciency) после трансплантации ESC (эмбриональных стволовых клеток) и, в случае HSC (гемопоэтических стволовых клеток), репопуляции кроветворной системы летально облученных мышей после трансплантации неочищенных клеток, полученных из костного мозга. Однако способность идентифицировать и изолировать стволовые клетки во многом зависит от использования различных маркеров, таких как поверхностные молекулы, факторы транскрипции и отток красителя. Для идентификации плюрипотентных клеток и стволовых клеток в определенных типах тканей доступны многочисленные маркеры, которые часто используются в комбинации. Хотя некоторые маркеры и тесты используются более универсально для распознавания стволовых клеток, особое внимание будет уделено тем, которые используются для идентификации стволовых клеток, полученных из костного мозга.
Кластер дифференциации (CD)34
CD34 — это гликопротеин клеточной поверхности, который традиционно используется для идентификации и очистки HSC и клеток-предшественников. Этот маркер, по-видимому, высококонсервативен среди видов млекопитающих. Экспериментальные данные свидетельствуют о том, что CD34 может участвовать в адгезии гемопоэтических клеток к стромальным клеткам в микросреде костного мозга. Однако совсем недавно были идентифицированы CD34-негативные HSC, называемые клетками боковой популяции (SP). Клетки SP считаются одними из самых примитивных HSC из-за их высокого пролиферативного потенциала и чрезвычайной эффективности в возвращении к местам кроветворения при инъекции мышам-реципиентам. Таким образом, экспрессия CD34 на HSC может быть связана со степенью активации этих клеток, причем CD34-негативные клетки являются наиболее примитивными и покоящимися.
Антиген стволовых клеток
Антиген стволовых клеток-1 (Sca-1) — это белок клеточной поверхности, часто используемый для идентификации мышиных HSC. Эта молекула может играть роль в определении линий???.
Отток красителя
Способность некоторых примитивных HSC выделять флуоресцентный краситель позволяет идентифицировать эту популяцию, называемую SP-клетками, с помощью проточной цитометрии. Эта способность, по-видимому, обусловлена увеличением количества или активности мембранных насосов (например, АТФ-связывающего кассетного транспортера [ABC-транспортера]), гипотеза, подтверждаемая обнаружением блокады оттока красителя препаратом верапамил, известным ингибитором этих оттокных насосов. Клетки SP не экспрессируют CD34 и описаны у многих видов. Как указывалось ранее, предполагается, что эти CD34-отрицательные клетки являются одними из самых примитивных HSC.
c - Kit
c - Kit — это трансмембранный тирозинкиназный рецептор, обнаруженный на HSC многих видов. Он связывает лигандный фактор стволовых клеток (SCF, также называемый фактором Steel) и играет важную роль в поддержании, пролиферации и дифференцировке HSC.
Lin
В дополнение к присутствию определенных маркеров (например, CD34, c - Kit, Sca - 1), отсутствие маркеров, присутствующих на дифференцированных клетках, использовалось для выделения и очистки HSC. Отрицательная по линии дифференцировки (Lin − ) классификация обычно означает, что клетки являются отрицательными по комбинации от 6 до 14 различных маркеров дифференцировки зрелых клеток крови.
Факторы транскрипции
Были идентифицированы факторы транскрипции, которые, по-видимому, играют важную роль в регуляции плюрипотентности стволовых клеток и их недифференцированного состояния. Наиболее примечательными являются факторы транскрипции Oct-4, Nanog и Sox-2, которые используются в качестве маркеров эмбриональных и взрослых стволовых клеток.
СТВОЛОВЫЕ КЛЕТКИ, ПОЛУЧЕННЫЕ ИЗ КОСТНОГО МОЗГА
БИОЛОГИЯ СТВОЛОВЫХ КЛЕТОК
Регуляция выживания и плюрипотентности
Ниша
Концепция ниши важна для обсуждения выживания и дифференциации стволовых клеток. Ниши — это локальные тканевые микросреды, функция которых — поддерживать, сохранять и регулировать стволовые клетки. Эти микроокружения обнаруживаются в различных тканях по всему организму, например, в области выпуклости волосяного фолликула, около основания крипт в желудочно-кишечном тракте и, в случае HSC, рядом с эндостом и синусоидами костного мозга. Регуляция стволовых клеток их нишей осуществляется посредством физического контакта и межклеточных взаимодействий с соседними клетками, а также выработки растворимых факторов. Данные также указывают на то, что стволовые клетки обладают способностью влиять на клеточные элементы своей ниши. Например, HSC мышей, подвергшихся острому гемопоэтическому стрессу, обладают повышенной способностью направлять мезенхимальные клетки костного мозга в сторону остеобластической дифференцировки в результате действия костных морфогенетических белков (BMP), полученных из HSC.
Молекулярные механизмы
Какие факторы заставляют стволовые клетки идти по пути самообновления и оставаться недифференцированными по сравнению с прогрессированием в сторону дифференциации линий и фенотипов зрелых клеток? Ответ на этот вопрос постоянно развивается по мере выяснения конкретных молекулярных механизмов. В эмбриональных стволовых клетках мышей выявлено несколько факторов, важных для поддержания выживания, самообновления и плюрипотентности стволовых клеток. Эти факторы делятся на внешние факторы (например, цитокины) и внутренние факторы (например, факторы транскрипции).
Фактор ингибирования лейкемии (LIF) LIF — это цитокин класса интерлейкина (IL) 6, который предотвращает дифференциацию эмбриональных стволовых клеток мышей в культуре. Связывание LIF с его мембранным рецептором приводит к активации множества сигнальных путей, таких как Янус-киназа/трансдукторы сигнала и активаторы транскрипции (JAK/STAT), фосфоинозитид-3-киназа (PI3K) и киназы, связанные с внеклеточным сигналом (ERK). Хотя эти пути являются общими нисходящими сигналами многих цитокинов, в данном контексте их активация, как правило, способствует поддержанию самообновления и плюрипотентности. Однако в этом примере активация ERK, по-видимому, способствует дифференцировке эмбриональных стволовых клеток мышей. Таким образом, LIF может активировать сигналы, которые либо способствуют, либо препятствуют поддержанию недифференцированного состояния, и именно баланс между этими нисходящими эффектами (обычно благоприятствующими самообновлению и плюрипотентности) определяет результат.
Костный морфогенетический белок 4 (BMP 4) и основной фактор роста фибробластов (FGF). BMP4 и основной FGF являются дополнительными примерами внешних факторов, которые способствуют самообновлению и плюрипотентности эмбриональных стволовых клеток мышей. В случае BMP4 он, по-видимому, работает в синергетическом состоянии с LIF. Вероятно открытие дополнительных сигнальных путей и факторов, поскольку факторы, важные для эмбриональных стволовых клеток мышей, не универсальны при применении к эмбриональным стволовым клеткам человека.
Wnt Были идентифицированы специфические внешние факторы, участвующие в самообновлении HSC. Сигнальный путь Wnt стимулирует самообновление HSC, одновременно подавляя дифференцировку HSC. Индукция активации β-катенина, нисходящего компонента сигнального пути Wnt, приводит к усилению самообновления мышиных HSC и ограничивает дифференциацию этих клеток. При добавлении ингибиторов пути Wnt к мышиным HSC и факторам роста пролиферация HSC подавляется.
Тирозинкиназа с доменами 2, подобными иммуноглобулину и эндотелиальному фактору роста (Tie2), и ангиопоэтин-1. Сигнализация Tie2/ангиопоэтина-1 также участвует в выживании HSC. Tie2 — это рецепторная тирозинкиназа, экспрессирующаяся на некоторых HSC. Ангиопоэтин-1 является лигандом рецептора Tie2 и способствует покою и повышению адгезии мышиных HSC к стромальным клеткам костного мозга. Считается, что регуляция состояния покоя и поддержание его в нише HSC важны для выживания HSC за счет защитного эффекта от миелосупрессивных стрессов.
Другие цитокины К другим цитокинам, важным для регуляции выживания HSC, относятся SCF, тромбопоэтин (TPO), BMP, FGF, инсулиноподобный фактор роста 2 (IGF-2) и интерлейкин (IL)-10. SCF и TPO являются общими компонентами большинства комбинаций цитокинов, используемых при культивировании и размножении HSC. Хотя TPO является основным цитокином, участвующим в образовании мегакариоцитов и тромбоцитов, было также показано, что он оказывает значительное воздействие на HSC. In vitro TPO способствует выживанию и размножению HSC, а у мышей, генетически измененных с целью отсутствия TPO или его рецептора, наблюдается значительно меньше стволовых клеток.
Факторы транскрипции Внутренние факторы, регулирующие недифференцированное состояние эмбриональных стволовых клеток, состоят в основном из факторов транскрипции. Наиболее примечательными являются Oct-4, Nanog и Sox-2. Эти факторы обнаружены в плюрипотентных клеточных линиях, и, как правило, их подавление приводит к дифференциации стволовых клеток. Присутствие Oct-4 и Sox-2, по-видимому, необходимо для плюрипотентности; Однако целевые гены для этих факторов транскрипции еще полностью не охарактеризованы. Также были описаны несколько факторов транскрипции и регуляторов клеточного цикла, управляющих самообновлением HSC.
Микрорибонуклеиновые кислоты (mi RNA) mi RNA представляют собой дополнительный внутренний молекулярный механизм, предположительно участвующий в поддержании плюрипотентных стволовых клеток. miRNA — это короткие одноцепочечные молекулы РНК, которые регулируют функцию генов, подавляя трансляцию посредством отжига, а иногда и деградации мРНК. Обнаружены новые miRNA, которые, по-видимому, преимущественно экспрессируются в недифференцированных эмбриональных стволовых клетках. Кроме того, оценка профилей экспрессии miRNA из эмбриональных стволовых клеток разной степени дифференцировки, а также из клеток зрелых тканей показывает подавление или потерю специфических микроРНК по мере перехода клеток в более дифференцированное состояние.
Регуляция дифференциацировки
Дифференциация стволовых клеток в определенные линии контролируется или направляется такими факторами, как цитокины, взаимодействие ниш и регуляторы самообновления и плюрипотентности. Цитокины влияют на определение линии стволовых клеток и направляют ее, и многие из них были описаны в контексте дифференцировки гемопоэтических стволовых клеток и клеток-предшественников (рис. 3.2; см. главы 6–11).
Стромальные клетки, составляющие нишу стволовых клеток, влияют на дифференциацию и определение линии посредством физического взаимодействия между клетками и выработки растворимых или связанных с клетками факторов (например, цитокинов). Наконец, как уже было описано ранее, существуют регуляторы, способствующие самообновлению и плюрипотентному состоянию стволовых клеток. Для того чтобы произошла дифференциация, эти регуляторы должны быть ингибированы или подавлены. С точки зрения механизма можно выделить две общие категории, по которым клетки ограничивают приверженность линии. Первый из них включает спектр поверхностных рецепторов, адгезионных белков и сигнальных путей, экспрессируемых данной клеткой. Например, цитокины играют важную роль в развитии клеточных линий. Однако если у стволовой или прогениторной клетки отсутствует рецептор цитокина, то этот цитокин не окажет практически никакого эффекта на свою цель. Подавление генов — это второй механизм, посредством которого может происходить ограничение генетической линии. Для того чтобы клетки дифференцировались, активируются или подавляются определенные гены, направляя клетки в сторону определенной линии. Это может быть достигнуто посредством таких механизмов, как метилирование ДНК и модификация гистонов, которые изменяют транскрипционное состояние хроматина.

ЗАБОЛЕВАНИЯ, СВЯЗАННЫЕ СО СТВОЛОВЫМИ КЛЕТКАМИ
Система кроветворения наглядно иллюстрирует последствия отказа стволовых клеток. HSC отвечают за постоянную замену всех клеточных компонентов крови; при недостаточности HSC развиваются цитопении (например, анемия, лейкопения, тромбоцитопения) и связанные с ними клинические проявления (например, летаргия, инфекция, кровотечение). Неудача HSC может быть результатом ряда основных патологических процессов, включая токсическое или лекарственно-опосредованное повреждение, иммуноопосредованное повреждение, инфекционные агенты (например, парвовирус, вирус лейкемии кошек и Ehrlichia spp.), недостаточную стимуляцию цитокинами и факторами роста, а также нарушение или повреждение ниши стволовых клеток (например, миелофтиз, ишемия, воспаление). (См. Раздел II.)
Стволовые клетки и пролиферативные заболевания
Подобно тому, как взрослые стволовые клетки отвечают за замену зрелых клеток и тканей, существуют убедительные доказательства того, что клетки со свойствами стволовых клеток лежат в основе патологии по крайней мере некоторых видов рака. Гипотеза о раковых стволовых клетках основана на нескольких основных наблюдениях. Первым из них является наблюдение за гетерогенностью опухоли. Многие опухоли состоят из клеток с различной морфологией и фенотипом, которые в некоторых случаях отдаленно напоминают исходную ткань. Это предполагает определенную степень дифференциации внутри популяции опухолевых клеток, что приводит к изменчивости структуры и функций. Более примитивная клетка-предшественник (т. е. раковая стволовая клетка) предположительно может дать начало различным фенотипам внутри опухоли. Второе наблюдение заключается в том, что трансплантация опухоли требует относительно большого количества раковых клеток, что свидетельствует о том, что только небольшое количество клеток в данной опухоли обладает способностью образовывать опухоль. Раковые стволовые клетки присутствуют в опухоли в небольшом количестве, поэтому для обеспечения присутствия этих клеток потребовалось бы сравнительно большое количество ткани. Как и обычные стволовые клетки, раковые стволовые клетки обладают основными функциональными свойствами самообновления и способностью к дифференциации.
У людей доказательства наличия раковых стволовых клеток были получены при раке кроветворной системы, головного мозга, молочной железы, толстой кишки, предстательной железы, костей и яичников, а также имеются некоторые доказательства существования раковых стволовых клеток у животных. Подтверждение существования раковых стволовых клеток у человека заключается в идентификации подгруппы опухолевых клеток, которые экспрессируют маркеры стволовых клеток и обладают исключительной или повышенной способностью образовывать опухоли in vitro или in vivo. Существуют данные, что раковые стволовые клетки могут возникать из нормальных стволовых клеток и/или клеток-предшественников, которые приобрели способность к самообновлению. Происхождение раковых стволовых клеток продолжает изучаться.
Существование раковых стволовых клеток имеет очевидные последствия для понимания биологии рака и лечения, по крайней мере, некоторых видов рака. Например, многие химиотерапевтические препараты воздействуют на быстро делящиеся клетки. Однако раковые стволовые клетки относительно медленно делятся, что позволяет им выживать при использовании традиционных методов лечения. Для эффективного лечения этих типов неоплазий будут важны новые терапевтические методы, направленные на устранение раковых стволовых клеток.
Сокращения и аббревиатуры
ОПРЕДЕЛЕНИЕ И ИСТОРИЯ
На сегодняшний день определено более 339 молекул CD. Этот взрывной рост числа молекул CD является результатом использования методов молекулярной биологии для идентификации новых молекул. Целью данной главы является предоставление наиболее актуального списка антигенов CD, распознаваемых различными антителами у животных ветеринарного направления. В таблице 4.1 обобщены современные знания о важных ветеринарных антигенах CD, включая клоны MAb, которые реагируют с антигенами CD собак, кошек, жвачных животных (крупного рогатого скота, овец и коз), свиней и лошадей; относительный(е) размер(ы) молекулы; топология в мембране; распределение в тканях; известная физиология; и видовая реактивность моноклональных антител. Включены избранные ключевые ссылки для каждого антигена CD.
Более подробная информация о молекулах CD, описанных в настоящее время, доступна на нескольких веб-сайтах, включая таксономическую ключевую программу, поддерживаемую Университетом штата Вашингтон, которая разработана для предоставления информации о конкретном городе моноклональных антител, специфичных для молекул лейкоцитарной дифференцировки внутри видов и между видами (http://www.vetmed.wsu.edu/tkp/). На этом сайте имеются ссылки на коммерческие компании, которые являются основными поставщиками ветеринарно-специфических моноклональных антител, включая Serotec (http://www.ab-direct.com/antibodies/_-500.html ) и Veterinary Medical Research & Development (VMRD). Международный обзор белков в Интернете (PROW) ( http://mpr.nci.nih.gov/prow/ ) предоставляет исчерпывающую базу данных с полным перечнем антигенов CD и ссылками на первичные последовательности нуклеиновых кислот и белков человеческих антигенов CD.

СТРУКТУРА И ИНТЕГРАЦИЯ МЕМБРАННЫХ АНТИГЕНОВ
ТРАНСМЕМБРАННЫЕ БЕЛКИ
Трансмембранные белки типов I и II имеют только одну трансмембранную α-спираль, содержащую 20–25 гидрофобных аминокислот. Белки типа I имеют N-концевую сигнальную последовательность эндоплазматического ретикулума (ER), которая расщепляется после того, как молекула попадает в ER. Это наиболее распространенный способ интеграции мембраны среди антигенов CD, перечисленных в таблице 4.1. Белок гликозилируется в аппарате Гольджи, если у белка есть сайт гликозилирования, а затем экспрессируется на поверхности клетки. Белки типа I закреплены в мембране таким образом, что их гидрофильный N-концевой участок находится на экзоплазматической стороне, а их гидрофильный C-концевой участок — на цитоплазматической стороне. Эти белки обычно представляют собой рецепторы и/или лиганды клеточной поверхности (например, CD4), многие из которых принадлежат к суперсемейству иммуноглобулинов (Ig).
Однопроходный трансмембранный белок типа II (II)
Трансмембранные белки типа II не имеют расщепляемой сигнальной последовательности ER и ориентированы таким образом, что их гидрофильный С-конец находится на экзоплазматической стороне, а гидрофильный N-конец — на цитоплазматической стороне. Эти белки имеют внутренний гидрофобный сигнал ER и мембранную якорную последовательность. Поскольку эти белки могут высвобождаться с поверхности клетки, они также могут действовать как плазменные белки, оказывая физиологическое воздействие на клетки, несущие контрлиганды. 1 CD13 — цинксвязывающая металлопротеаза, которая облегчает презентацию антигена путем отщепления N-концевых аминокислот от пептидов, связанных с MHC класса II, — является примером белка типа II.
Многопроходный трансмембранный белок (III)
Трансмембранные белки типа III пересекают мембрану многократно (2, 3, 4, 5, 7 или 12) раз и также называются тетраспанинами. Наиболее часто встречающиеся тетраспанины включают те, которые пронизывают мембрану четыре и семь раз. Структурные исследования показывают, что трансмембранные области представляют собой альфа-спирали. Если белок типа III имеет четное число трансмембранных альфа-спиралей, его N- и C-концы ориентированы к одной и той же стороне мембраны. Многие из этих молекул клеточной поверхности функционируют как рецепторы для растворимых молекул, таких как простагландины и хемокины. Примерами семи трансмембранных белков типа III являются рецептор интерлейкина-8 (ИЛ-8) (CD128) и рецептор C5a (CD88). CD9 является примером четырехпроходного трансмембранного белка. 1 CD47 — пример белка типа III с пятью трансмембранными последовательностями. Транспортный белок множественной лекарственной устойчивости MDR-1 (CD243) имеет 12 трансмембранных участков.
Трансмембранный белок Gpi (V)
Трансмембранные белки типа V используют GPI-якоря, прикрепленные к C-концевому остатку белка. Молекулы с GPI-якорем имеют на N- и C-конце последовательность сигнала секреции, которая расщепляется и заменяется GPI-якорем после синтеза молекулы и входа в ER. Примером GPI-связанного гликопротеина является поверхностный белок CD14, участвующий в очистке организма от грамотрицательных патогенов, связанный с белком, связывающим липидный полисахарид (рис. 4.1). Другими примерами GPI-связанных гликопротеинов являются CD56 и CD90.
Этот маркер часто используется для дифференциации острых незрелых лейкозов лимфоидного или миелоидного происхождения от хронического лимфолейкоза или лейкемических стадий лимфомы. К поверхностным антигенам С-клеток, специфичным для определенной линии и полезным для определения линий лейкоцитарных клеток, относятся CD3, который экспрессируется исключительно на зрелых Т-клетках, а также CD79 α и β, которые являются частью рецептора поверхностного антигена В-клеток. К поверхностным антигенам, не связанным с рестрикцией линий, относятся такие антигены, как CD45, который обнаруживается во всех линиях лейкоцитов (миелоидных и лимфоидных). Хотя последнее исследование CLAW проводилось в 1993 году, оно стало важным совместным проектом, поскольку у собак была выявлена аномальная экспрессия антигена CD4, у которых высокая плотность CD4 наблюдается на нейтрофилах в дополнение к Т-хелперам. 34 Многие другие различия в экспрессии антигена CD являются уникальными среди домашних животных по сравнению с людьми (например, высокая экспрессия рецептора γ δ T-клеток у свиней и жвачных животных5,36 и совместная экспрессия CD4 и CD8 на зрелых T-клетках у свиней).

Сокращения и аббревиатуры
Гемопоэтические стволовые клетки
Приверженность линии обычно следует за экспрессией факторов транскрипции, ограничивающих линию (таблица 5.1). Индукция этих факторов транскрипции происходит посредством комбинации специфических взаимодействий рецепторов и лигандов цитокинов, менее специфических путей передачи сигнала и наложения высокоспецифичных, пермиссивных влияний микросреды со стороны стромальных клеток, эндотелиальных клеток, адипоцитов, остеобластов, белков ECM, адгезивных цитокинов и трабекулярной кости.

На основе экспериментов in vivo на мышах сложилась несколько запутанная номенклатура, посвященная ранним детерминированным гемопоэтическим клеткам-предшественникам. Колонии в селезенке, которые появляются у летально облученных мышей, восстановленных с помощью HSC, называются колониеобразующими клетками в селезенке (CFC-Ss), которые имеют ограниченную способность к самообновлению. Неизвестно, встречаются ли подобные клетки у других видов, поскольку CFC-S определяются специфическими экспериментальными условиями на мышах.

Гематопоэтические предшественники и прекурсоры
В правой части рисунка 5.1 клонигенные типы клеток предположительно не обладают способностью к самогенерации. Однако зрелые типы клеток, такие как макрофаги и тучные клетки, могут обладать существенными свойствами самогенерации. Первоначальная модель кроветворения не включала в себя понятие пластичности. В современной модели концепция пластичности предполагает, что точность линии и прогрессивное ограничение пролиферативной способности в традиционной модели не являются абсолютными. Пластичность рассматривается как свойство как пролиферации, так и приверженности линии. То, как регулируется пластичность в физиологических и патологических условиях, изучено недостаточно. Однако ясно, что ни HSC, ни любой из олигопотентных предшественников не обладают неограниченной способностью к самовоспроизведению, которой обладают эмбриональные стволовые клетки.
Видовая специфичность гемопоэза
Между кроветворением человека и мышей существуют определенные существенные различия. Человеческие HSC экспрессируют Flt3, тирозинкиназный рецептор для лиганда FLT3, тогда как мышиные HSC этого не делают. Однако общие предшественники лимфоцитов (CLP) как у мышей, так и у человека экспрессируют Flt3. Такие различия важны, поскольку некоторые острые миелоидные лейкозы человека связаны с конститутивной активацией Flt3. Потенциальная полезность терапии препаратами, ингибирующими Flt3-киназу, зависит от того, проявят ли другие виды подобные мутации. Хотя иерархические отношения, наблюдаемые в кроветворении мышей, в целом сохраняются и у других видов (рис. 5.1), экспрессия ряда поверхностных антигенов на каждой стадии развития у мышей и людей различна, и эти и другие различия могут встречаться и у других видов. Однако были также выявлены и важные сходства. На основании последних данных молекулярно-генетического понимания циклической нейтропении у серых колли и циклического кроветворения у людей некоторые особенности кроветворения кажутся схожими. Например, аллометрическое масштабирование правильно предсказывает гранулопоэтическую цикличность у мышей (3 дня), собак (14 дней), людей (19–21 день) и слонов (60 дней).
МИКРООКРУЖЕНИЕ КОСТНОГО МОЗГА
Лучше всего охарактеризованным интегрином на ГСК является очень поздний антиген 4 (VLA - 4 или α 4 β 1), который связывается с фибронектином в ECM, а также с молекулой адгезии сосудистой клетки 1 (VCAM - 1) на соседних стромальных клетках. Связывание VLA - 4 с фибронектином опосредует адгезию ГСК и предшественников к микроокружению, хоуминг циркулирующих ГСК и предшественников в определенные области в костном мозге и передачу сигнала. Rho-гуанозинтрифосфатазы (ГТФазы), Ras-связанный субстрат 1 ботулинического токсина C3 (Rac1) и Rac2 также являются ключевыми регуляторами адгезии и миграции клеток в кроветворном микроокружении.
Микросреда костного мозга состоит из высокоспециализированных и интегрированных микроанатомических функциональных единиц, называемых нишами. Две наиболее изученные гемопоэтические ниши – это ниша ГСК и ниша эритробластических островков (рис. 5.2 и 5.3).
Ниша гемопоэтических стволовых клеток
Выживание, пролиферация и дифференцировка ГСК зависят от их пространственных и функциональных взаимоотношений с клетками и внеклеточным матриксом микроокружения. ГСК обогащаются в участках, прилегающих к эндостальной поверхности кости. Специализированная подгруппа активированных остеобластов демонстрирует лиганды и рецепторы, которые облегчают хоуминг и временную стыковку ГСК и регулируют медленную цикличность или быструю мобилизацию ГСК по мере необходимости. Взаимодействие между стромальным фактором 1 (SDF-1 или CXCL12), вырабатываемым остеобластами, и хемокиновым рецептором CXC 4 (CXCR4), его родственным хемокиновым рецептором на ГСК, является центральным для этой ниши ГСК. SDF-1 рекрутирует покоящихся предшественников, участвует в их цикличности и выживании и сенсибилизирует их к дальнейшему синергетическому действию цитокинов, тем самым способствуя гемопоэтическому гомеостазу как в физиологических, так и в стрессовых условиях. Функция ГСК также регулируется остеобластами через паратиреоидный гормон (ПТГ), сигнальный путь Notch и взаимодействие ангиопоэтина 1 (Ang1) и его тирозинкиназы с рецептором иммуноглобулин-подобного и EGF-подобного доменов 2 (Tie2). Нарушение этой ниши HSC может привести к аномальной мобилизации клеток и способствовать экстрамедуллярной инфильтрации при лейкемии.


Эритропоэз происходит в отдельных нишах, называемых эритробластическими островками, которые состоят из центрального макрофага, окруженного кольцом развивающихся эритробластов, в костном мозге, печени плода и селезенке, а также в долгосрочных культурах костного мозга. Макрофаг передает важные сигналы развивающимся рубрибластам, фагоцитирует изгоняемые ядра метарубрицитов и переносит железо развивающимся рубрибластам. В отличие от мегакариоцитов, которые локализуются исключительно в синусоидах костного мозга, эритробластические островки расположены по всему костному мозгу.
Молекулы адгезии, опосредующие важные структурные и функциональные взаимодействия между развивающимися эритроидными клетками и центральными макрофагами, включают VCAM - 1/VLA - 4, α4 β1/VLA - 4, молекулу межклеточной адгезии - 4 (ICAM - 4)/αv и Е- кадгерин. Например, предполагается, что ICAM - 4 позволяет ретикулоцитам отделяться от центральных макрофагов, позволяя им проникать в кровоток. Ламинин и фибронектин и их рецепторы, экспрессируемые на поздних стадиях рубрибластов, также являются ключевыми компонентами дифференцировки ретикулоцитов. VLA - 4 и VLA - 5 участвуют в связывании эритроидных разрывообразующих единиц (BFU - Es) с гемопоэтическими стромальными клетками. Экспрессия этих молекул адгезии наиболее высока в КОЕ и эритроидных колониеобразующих единицах (КОЕ - Es) и постепенно снижается в процессе созревания эритроидов. Гемонектин и коллаген I типа поддерживают связывание BFU - Es in vitro, и роль тенасцина - С была доказана у мышей с нокаутом. Гемонектин отсутствует у анемичных мышей с лигандом Kit или дефицитом c - kit.
Кластер дифференцировки поверхностного антигена 44 (CD44) высоко экспрессируется почти на всех гемопоэтических клетках костного мозга и отвечает за взаимодействие этих клеток с коллагеном I и IV типов, фибронектином и гиалуронатом. Ретикулоциты экспрессируют лишь низкие уровни нескольких поверхностных рецепторов адгезии, таких как CD36 (рецептор тромбоспондина) и VLA - 4. Тромбоспондин служит адгезивным лигандом для коммитированных предшественников, включая колониеобразующие единицы - гранулоциты, эритроиды, моноциты, мегакариоциты (КОЕ - GEMM) и BFU - E. Взаимодействие молекул адгезии ретикулоцитов со стромальными клетками костного мозга и ВКМ может способствовать их проникновению в синусы костного мозга. Зрелые эритроциты (RBC) в нормальных условиях не экспрессируют молекулы адгезии.
ЦИТОКИНЫ И ЦИТОКИНОВАЯ СИГНАЛИЗАЦИЯ В КРОВЕТВОРЕНИИ
Большинство рецепторов гемопоэтических цитокинов представляют собой комплексы из нескольких субъединиц, за исключением тех, которые передают сигнал по одной цепи, таких как эритропоэтин (EPO), гранулоцитарно-макрофагальный колониестимулирующий фактор (GM - CSF) и тромбопоэтин (TPO). Гемопоэтические цитокины имеют длину около 200 аминокислот и содержат консервативную последовательность триптофан - серин - Х - триптофан - серин (W - S - X - W - S) во внеклеточном домене, который функционирует как часть лиганд- связывающего домена.
Цитокиновые рецепторы содержат на своих цитоплазматических концах стыковочные участки для тирозинкиназ януса (JAK1, JAK2, JAK3, TYK2), которые, присоединяясь к лиганд -связанной форме цитокинового рецептора, рекрутируют JAK, которые затем аутофосфорилируются. JAK-киназа затем фосфорилирует остатки тирозина на специфическом белке-трансдюсере сигнала и активаторе транскрипции (STAT), которым для большинства гемопоэтических цитокиновых рецепторов является STAT1, 3 или 5. JAK содержат каталитический домен Януса (JH) 1 и каталитически подобный, но неактивный домен JH2, критически важный для способности JAK саморегулироваться и опосредовать цитокин-индуцированные ответы. Активация STAT киназой Src также важна для пролиферации миелоидных клеток.
В нестимулированных клетках STAT присутствуют в виде цитоплазматических мономеров в нефосфорилированном состоянии. Фосфорилирование JAK-киназами приводит к димеризации за счет взаимных взаимодействий SH2-доменов с остатками фосфотирозина и, таким образом, активации STAT, который затем перемещается в ядро. STAT - это факторы транскрипции, которые предотвращают апоптоз или положительно регулируют выживаемость генов поздних клеток-предшественников и ранних клеток-предшественниц. Хотя существует множество генов, реагирующих на STAT, во многих различных типах клеток, регуляция STAT гемопоэтических предшественников осуществляется ограниченным типом клеток образом.
Клетки, формирующие колонии, реагируют на цитокины абсолютно линейно-ограниченным образом. Специфичность гемопоэтических цитокинов определяется экспрессией их соответствующих рецепторов клетками-предшественниками. Например, рубробласты экспрессируют рецепторы EPO, а миелобласты – нет, что отличается от относительно неравномерной экспрессии путей JAK/STAT. Учитывая важность фосфорилирования JAK в стимуляции пролиферации гемопоэтических предшественников, неудивительно, что мутации, приводящие к конститутивно активной JAK2, приводят к миелопролиферативным заболеваниям и лейкемии. Помимо важной роли взаимодействий JAK-STAT в гемопоэзе, также была показана функциональная вовлеченность путей Ras и фосфоинозитол-3-киназы (PI3K) после стимуляции культур костного мозга интерлейкином (IL)-3/IL-5 и GM-CSF in vitro.
ТАБЛИЦА 5.2 Гемопоэтические цитокины а
Супрессоры сигнализации цитокинов (SOCS) представляют собой семейство белков, которые регулируют силу и продолжительность каскада сигнализации, управляемого гемопоэтическими цитокинами. Они транскрипционно индуцируются сигнализацией JAK - STAT. Белки SOCS содержат домены гомологии src (SH) 2 и SOCS - бокс, которые опосредуют связывание с рецепторами цитокинов и связанными с ними JAK, а также напрямую ослабляют передачу сигнала. В дополнение к их транскрипционной индукции гемопоэтическими цитокинами и последующей самоограничивающейся стимуляции гемопоэза, другие цитокины, включая фактор некроза опухоли - α (TNF - α ), ИЛ - 1 и лиганды Toll - подобных рецепторов (например, липополисахарид), также индуцируют экспрессию SOCS, обеспечивая негативную регуляцию сигнализации гранулоцитарного колониестимулирующего фактора G - CSF).
Другой уровень регуляции обеспечивается фосфатазами. Передача сигналов и последующее кроветворение, вызванные фосфорилированными димерами STAT-белков, прекращаются путем удаления фосфатов из STAT-тирозинов тремя специфическими белковыми тирозинфосфатазами.
Трансформирующий фактор роста β (TGF - β), возможно, является наиболее мощным эндогенным негативным регулятором кроветворения. TGF - β подавляет экспрессию рецептора SCF, реакцию предшественников на SCF и прогрессирование клеточного цикла предшественников. Экспрессия рецепторов к TGF - β на примитивных гемопоэтических предшественниках и последующих стадиях созревания свидетельствует о широкой роли этого цитокина. Негативная регуляция эритропоэза с помощью TNF - α, связанного с TNF лиганда, индуцирующего апоптоз (TRAIL), IL - 6 и TGF - β, происходит, когда хроническое воспалительное заболевание увеличивает системную и местную концентрацию этих цитокинов в костном мозге.
Общие сведения


ОПРЕДЕЛЕНИЯ
ПРЕВРАЩЕНИЕ СТВОЛОВОЙ КЛЕТКИ В ЭРИТРОЦИТ
HSC мультипотентны и могут делиться на копии самих себя или на клетки, преданные определенным линиям клеток крови. Мультипотентная HSC (рис. 6.1) сначала дифференцируется в общий миелоидный предшественник (CMP), который затем способен дифференцироваться в любые клетки крови, кроме лимфоцитов. CMP имеет ограниченную способность к самообновлению и, в зависимости от сигналов окружающей среды, способен дифференцироваться в предшественника гранулоцитов/макрофагов или предшественника мегакариоцитов/эритроидов (MEP). CMP и MEP у людей и мышей можно идентифицировать по их специфическому фенотипу клеточной поверхности. Дифференциация от CMP к MEP связана с экспрессией различных ген-специфических факторов транскрипции, рецепторов факторов роста и функциональных белков и зависит от множества факторов роста. Ген-специфические факторы транскрипции являются преимущественно активаторами транскрипции и обычно функционируют как комплекс белков, которые связываются со специфическими последовательностями дезоксирибонуклеиновой кислоты (ДНК).
MEP дифференцируются в единицы, формирующие взрыв - эритроидные клетки (BFU - Es). Хотя BFU - Es запрограммированы на дифференциацию только в эритроидные клетки, микроскопически они напоминают лимфоциты. Эти клетки идентифицируются по их росту от одной клетки до нескольких тысяч клеток в культуре. Колониеобразующая единица - эритроид (CFU - E) дифференцируется из BFU - E. В культуре каждая CFU - E делится на 8 - 32 клетки. Таким образом, BFU - Es имеют более высокий пролиферативный потенциал, чем более дифференцированные CFU - E. Эти клетки-предшественники дифференцируются в клетки-предшественники, которые могут быть распознаны как эритроидные клетки на основании морфологических характеристик. Самым ранним предшественником является рубрибласт (рис. 6.2). Это крупная клетка с глубоко базофильной цитоплазмой; круглым, центрально расположенным ядром; мелкозернистым, но глубоко окрашивающимся ядерным хроматином; и одним или несколькими заметными ядрышками. Глубоко базофильная цитоплазма помогает отличить ранние эритроидные от ранних миелоидных предшественников. Рубрибласты проходят ряд стадий дифференцировки, которые приводят к прогрессивному уменьшению размера клетки, постепенному увеличению концентрации цитоплазматического гемоглобина и постепенной конденсации ядерного хроматина. Прорубрицит похож на рубрибласт, за исключением того, что он не содержит ядрышка. Последующие стадии называются базофильными рубрицитами, полихроматофильными рубрицитами и метарубрицитами. Эти клетки имеют увеличивающееся количество гемоглобина, что влияет на качество окрашивания цитоплазмы. Базофильные рубрициты имеют самую низкую концентрацию гемоглобина и, следовательно, все еще имеют глубоко базофильную цитоплазму. Полихроматофильные рубрициты имеют цитоплазму, которая варьируется от сине-серого до серого, в зависимости от концентрации гемоглобина. Метарубрициты имеют наибольшее количество гемоглобина и, следовательно, имеют серую или красноватую цитоплазму. Метарубрициты также имеют плотный, гомогенный ядерный хроматин. На этом этапе ядро выталкивается, образуя полихроматофильные эритроциты, которые окрашиваются как ретикулоциты прижизненными красителями, такими как новый метиленовый синий. Все предшественники, за исключением метарубрицитов, способны к делению.
Номенклатура предшественников эритроидов у человека и мышей отличается от той, которая используется в ветеринарии. Самый ранний предшественник эритроидов называется проэритробластом, а последующие клетки называются базофильными эритробластами, полихроматофильными эритробластами и ортохроматическими эритробластами (таблица 6.1).
Развитие и созревание ретикулоцитов
Изгнание ядер из метарубрицитов связано с изменениями в промежуточных филаментах и микротрубочках, которые приводят к перестройке мембранного цитоскелета. Вытесненные ядра быстро поглощаются макрофагами костного мозга. Затем ретикулоциты покидают костный мозг, хотя точный механизм неизвестен. Ретикулоциты содержат остаточные митохондрии, мембраны аппарата Гольджи, рибосомы и микротрубочковые компоненты и могут иметь примерно на 35% больший объем, чем зрелые эритроциты. Во время созревания в кровотоке ретикулоциты приобретают гемоглобин и теряют органеллы, площадь поверхности мембраны, объем и многочисленные белки клеточной поверхности. Белки клеточной поверхности теряются в виде экзосом или мембранных пузырьков, которые содержат белки, специально отобранные для удаления.
Стрессовые ретикулоциты высвобождаются из костного мозга во время эритропоэтического стресса. Стрессовые ретикулоциты крупнее и менее зрелые, чем нормальные ретикулоциты, и содержат больше митохондрий, рибосом и других органелл. Их мембраны более механически жесткие и нестабильные, и они содержат адгезивные белки, которые обычно теряются перед высвобождением.
Молекулы клеточной поверхности на развивающихся эритроидных клетках
Эритроидные клетки экспрессируют различные молекулы на поверхности клеток, некоторые из которых регулируются в процессе развития. Краткое описание важных молекул на поверхности эритроидных клеток представлено в таблице 6.2.
Ретикулоциты в ветеринарии
Количество ретикулоцитов, циркулирующих в норме, варьируется у разных видов. У некоторых видов ретикулоциты присутствуют в норме (собаки, кошки, грызуны, кролики, свиньи, морские свинки), тогда как у других (коровы, козы, овцы) ретикулоциты присутствуют только в период регенеративной реакции. У лошадей, за редкими исключениями, ретикулоциты отсутствуют даже в период регенеративной реакции.
Физиологический процесс
A. Прекурсоры эритроцитов
1. Эритропоэз, производство эритроцитов, является частью кроветворения, сложной системы, включающей стволовые клетки и цитокины. Несколько цитокинов взаимодействуют с Epo (эритропоэтином), стимулируя репликацию и дифференцировку клеток-предшественников в стволовые клетки, которые реагируют на Epo либо делением, либо дифференцировкой в рубрибласты. Эпо также способствует эритропоэзу, ингибируя апоптоз.
2. Эритропоэтин вырабатывается в основном печенью плода и почками взрослого животного. Интерстициальные клетки почек вырабатывают эритропоэтин в ответ на почечную гипоксию, которая может быть вызвана анемией, плохой оксигенацией крови (например, из-за большой высоты или легочных заболеваний) или плохой почечной перфузией.
a. Гипоксия позволяет индуцируемому гипоксией фактору 2α (HIF-2α) транслоцироваться в ядро, связываться с HIF-1β и, таким образом, активировать транскрипцию гена эритропоэтина.
b. В здоровом состоянии меньшее количество HIF-2α достигает ядра, что способствует выработке эритропоэтина, поскольку при более высоком уровне O2 HIF-2α остается гидроксилированным ферментами пролилгидроксилазного домена (PDH), что позволяет ему связываться с белком-супрессором опухоли фон Гиппеля Линдау (VHL) и подвергаться ферментативной деградации. Мутации в генах белков PDH, HIF или VHL могут нарушать регуляцию эритропоэза и вызывать эритроцитоз.
3. После того, как клетка-предшественница (КОЕ-Е) дифференцируется в рубрибласт, происходят два основных клеточных процесса:
a. Клетки подвергаются митозу, в результате чего образуются новые и более мелкие клетки. Имеющиеся данные свидетельствуют о том, что увеличение [гемоглобина, Hgb] в развивающихся клетках снижает синтез ДНК и в конечном итоге ограничивает митоз.
b. Клетки продуцируют информационную РНК для синтеза Hgb и белков цитоскелета.
4. Эритроциты развиваются в ассоциации с макрофагами (кормящими клетками), которые, как полагают, обеспечивают сигналы, факторы роста и, вероятно, железо эритроидному предшественнику для синтеза Hgb. На более поздних стадиях эритропоэза у млекопитающих ядра nRBC (нормобласт) вытягиваются и поглощаются макрофагами. Образующаяся безъядерная клетка называется ретикулоцитом, если она содержит
окрашиваемую РНК.
5. Когда nRBC теряет свое ядро и способность продуцировать мРНК для синтеза белка, он вскоре теряет способность продуцировать Hgb и ферменты. Активность ферментов RBC наиболее высока в ретикулоцитах и медленно снижается с возрастом. Митохондрии разрушаются, так что гликолиз становится путем производства АТФ (за исключением свиней, которые используют альтернативный энергетический путь с участием инозина).
B. Эритроциты в крови
1. В отличие от других домашних млекопитающих, эритроциты верблюдообразных представляют собой тонкие эллипсоидные клетки, содержащие большую концентрацию Hgb. Их тонкость способствует газообмену, которому у других видов способствует двояковыпуклость эритроцитов и связанное с этим высокое отношение площади поверхности к объему. Они обладают пониженной деформируемостью, что может быть связано с другим составом мембран и взаимодействием с цитоскелетом, но они могут значительно набухать и поэтому устойчивы к гипотоническому лизису, который в противном случае может возникнуть при регидратации после периодов обезвоживания.
2. Разрушение эритроцитов в организме (старение)
а. В час погибает около 200-250 миллионов эритроцитов на килограмм массы тела!
b. С возрастом активность ферментов эритроцитов снижается, они накапливают окислительные повреждения, теряют мембрану в результате образования пузырьков и становятся более жесткими, сфероидальными и менее способными проходить через поры селезеночной щели. Появляются мембранные антигены (например, на кластерах белка band 3), и они связываются природными антителами, которые активируют комплемент. Эта опсонизация и другие изменения мембран помогают макрофагам осуществлять фагоцитоз стареющих эритроцитов.
C. Эритроциты в селезенке
1. Селезенка собак, лошадей и крупного рогатого скота имеет синусоиды и красную мякоть, которые полны эритроцитов; 50-60 % эритроцитов лошади может находиться в ее селезенке. Поврежденные или менее деформируемые эритроциты удаляются макрофагами, которые расположены рядом с синусоидами или присутствуют в красной пульпе. При сокращении селезенки эритроциты попадают в системный кровоток.
2. В селезенке кошек отсутствуют пазухи и обеспечивается открытое кровообращение без артериовенозных соединений, которые наблюдаются у собак. Эритроциты, по-видимому, попадают из красной пульпы в венозный кровоток через открытые венулы пульпы, а не через узкие межэндотелиальные синусовые щели, которые у других видов важны для удаления (“изъязвления”) включений эритроцитов, таких как тельца Хауэлла–Джолли, тельца Хайнца, сидерозные гранулы и внутриклеточные организмы. Эти прорези также способствуют удалению плохо деформируемых эритроцитов. Менее ограниченное прохождение эритроцитов через селезенку кошек может также снижать ремоделирование ретикулоцитов и объяснять длительный неретикулоцитарный макроцитоз, описанный у кошек после регенеративной анемии.
3. Большинство метарубрицитов и ретикулоцитов, которые ежедневно выделяются из костного мозга, временно задерживаются в селезенке млекопитающих.
II. Кинетика эритроцитов
A. Уровень эритроцитов в крови определяется по относительной скорости образования эритроцитов, перемещению эритроцитов в синусы селезенки и из них и разрушению эритроцитов.
B. Выработка эритроцитов зависит от степени и продолжительности воздействия эритропоэтина и способности клеток-предшественников реагировать на эритропоэтина.
III. Структура, функции, синтез и деградация Hgb
A. Структура гемоглобина.
1. Hgb представляет собой тетрамер, в котором каждый глобин (полипептидная цепь аминокислот) связан с отдельным гемом, который связывает O2; его относительная молекулярная масса составляет около 64 000. У взрослых здоровых млекопитающих каждая молекула Hgb содержит две α-цепи и две β-цепи глобинов. Примерно 95 % эритроцитов в пересчете на сухую массу составляют Hgb, в каждом эритроците содержится от десятков до сотен миллионов молекул Hgb.
а. Скорости синтеза гема и глобина сбалансированы и регулируются друг другом. Если в предшественнике содержится очень мало гема и доступен Fe2+, синтез гема и глобина должен увеличиться.
b. Если синтез глобиновой цепи снижен, у животного развивается талассемия.
c. Если генные мутации приводят к появлению аномальных аминокислотных последовательностей в глобинах, у животного развивается гемоглобинопатия.
2. Железо в геме находится в двухвалентном состоянии (Fe2+) и поддерживается в этом восстановленном состоянии ферментативными реакциями, катализируемыми Cb5R (NADH-метгемоглобинредуктазой, обычно называемой метгемоглобинредуктазой) и NADPH-диафоразой (NADPH-
метгемоглобинредуктазой).
3. Аминокислоты в цепях глобина поддерживаются в восстановленном состоянии с помощью восстановительных реакций с участием GR и каталазы.
B. Функция Hgb (гемоглобина).
1. Hgb переносит O2 из легких в ткани. В здоровом состоянии артериальная кровь почти на 100% насыщена O2. Метилгидроксид (с Fe3+) не переносит O2.
2. Гемоглобин играет две основные роли в переносе CO2 из тканей в легкие:
а. Когда CO2 диффундирует в эритроциты, карбоангидраза катализирует его реакцию с H2O с образованием H+ и HCO3−. Hgb действует как буфер (H+ + Hgb− → HHgb) для удаления H+, а HCO3− диффундирует из клеток в плазму. Буферизация H+ с помощью Hgb способствует дополнительному преобразованию CO2 в HCO3−. Когда эритроциты возвращаются в легкие, реакция меняется на противоположную, и при выдохе выделяется CO2. Около 70 % CO2, образующегося в тканях, транспортируется в легкие через эту систему.
b. Когда CO2 диффундирует в эритроциты, часть его связывается с Hgb, образуя карбаминогемоглобин. Около 20 % CO2, образующегося в тканях, транспортируется в легкие через эту систему.
C. Синтез гемоглобина.
1. Это происходит в предшественниках эритроцитов (рубрибластах и ретикулоцитах) в результате ряда реакций (рис. 3.2).
2. Свинец ингибирует 5-аминолевулинат-синтазу, порфобилиноген-синтазу, феррохелатазу и копропорфириногеноксидазу.
а. При отравлении свинцом ингибирование этих ферментов приводит к повышению концентрации предшественников гема в эритроцитах. В совокупности порфобилиноген и протопорфирин IX называются порфиринами.
b. Порфирия - это состояние, при котором повышаются концентрации порфиринов в эритроцитах, плазме или моче. Это может быть приобретенное заболевание, например, при отравлении свинцом, или врожденное. Животные с порфирией склонны к фоточувствительности.
3. Скорость созревания предшественников эритроцитов зависит от цитоплазматического уровня гемоглобина. Если синтез Hgb неполон, дополнительные митозы в процессе развития клетки приводят к образованию эритроцитов меньшего размера.
D. Тип Hgb, синтезируемого у человека, может варьироваться в зависимости от возраста и состояния здоровья животного. У людей существует три основных типа Hgb: (i) эмбриональный Hgb, содержащий в основном два α-глобина и два ε-глобина, (ii) фетальный Hgb, содержащий два α-глобина и два γ-глобина, и (iii) взрослый Hgb, содержащий в основном два α-глобина и два β-глобина. Эмбриональный Hgb заменяется взрослым Hgb во время внутриутробного развития у собак, кошек и лошадей; у этих животных эмбриональный Hgb отсутствует. Считается, что новорожденные жвачные животные имеют смесь Hgb плода (с двумя α-глобинами и двумя βF-глобинами) и взрослого Hgb, но только взрослый Hgb через несколько месяцев после рождения содержит два α-глобина и два βA-глобина.
E. Распад Hgb и метаболизм билирубина: После гибели эритроцитов в макрофагах селезенки, печени или костного мозга гемовая часть Hgb расщепляется в макрофагах до биливердина, а затем до Bu, при этом гемовый Fe2+ подвергается рециркуляции, и из каждой гемовой группы образуется одна молекула оксида углерода. Глобиновая часть Hgb расщепляется до аминокислот, которые также могут быть рециркулированы. Bu конъюгируется с Bc, который выводится из организма (рисунок 3.3). Угарный газ диффундирует в плазму, а затем в эритроциты, где образует карбоксигемоглобин; позже он высвобождается в легких и выдыхается. Из-за этого метаболизма карбоксигемоглобинемия часто встречается при гемолитической анемии и может сопровождаться внутренним кровотечением.

Рисунок 3.1. Кинетика эритроцитов в норме. Эритрон содержит три основных пула: (i) предшественники эритроцитов (в основном в костном мозге), (ii) эритроциты крови и (iii) эритроциты селезенки. После стимуляции Эпо колониеобразующие единичные эритроидные клетки (КОЕ-Е) дифференцируются в рубрибласты (Rb), а предшественники пролиферируют (посредством митоза) и созревают до образования эритроцитов (Es). Упорядоченный процесс созревания приводит к некоторому пирамидальному распределению популяций эритроидных клеток (показана только верхняя половина). После попадания в кровь эритроциты циркулируют по сосудистой системе, транспортируя O2 к тканям и CO2 в легкие. У большинства млекопитающих запасы эритроцитов находятся в селезенке. Стареющие эритроциты разрушаются макрофагами селезенки, образуя Bu из Hgb. Mt - метарубрицит; Pr - прорубрицит; Rc - рубрицит; и Rt - ретикулоцит.
Bu - неконъюгированный билирубин.
Hgb - гемоглобин.

Рисунок 3.2. Синтез гемоглобина у взрослых домашних млекопитающих.
• Синтез Hgb в предшественниках эритроцитов происходит в митохондриях и включает восемь ферментативных реакций, в которых образуются отдельные пиррольные кольца (этапы 1 и 2), полимеризуются в тетрапиррольные кольца (этапы 3 и 4) и модифицируются (этапы 5–7 )перед включением Fe2+ в протопорфирин IX с образованием гема, который покидает митохондрию. Каждая из четырёх молекул гема затем связывается с одной из двух α- или двух β-субъединиц белка, образуя тетрамерный Hgb с четырьмя центрами связывания O2; это может включать окисленную (Fe3+) форму гема (ферригем, гемин) с образованием MetHgb (не показано), который восстанавливается до Hgb. Реакция 1 является лимитирующей стадией синтеза гема; Для этого требуется витамин B6 (пиридоксин), и его активность подавляется снижением доступности железа и повышением концентрации гема в клетках.
• Наследственный или приобретенный дефицит одного или нескольких из восьми ферментов, участвующих в синтезе гемоглобина, вызывает порфирию. Ферменты для каждой реакции следующие: 1. 5-аминолевулинатсинтаза, 2. порфобилиногенсинтаза, 3. гидроксиметилбилансинтаза, 4. уропорфириноген III-синтаза, 5. уропорфириногендекарбоксилаза, 5. копропорфириногеноксидаза, 6. протопорфириногеноксидаза и 6. феррохелатаза.
δ-АЛК — дельта-аминолевулиновая кислота; сукцинил-КоА — сукцинил-коэнзим А

Рисунок 3.3. Физиологические процессы, связанные с билирубином.
- Разрушение эритроцитов внутри макрофагов селезенки (показано здесь), печени или костного мозга сопровождается деградацией гема и его превращением в Bu (Bilirubin unconjugated / билирубин неконъюгированный). Клинически незначительные количества Bu образуются при деградации гема, связанной с неэффективным эритропоэзом, и деградации других гем-содержащих молекул (каталазы, пероксидазы и цитохромов). Когда Bu покидает макрофаг, он образует нековалентную связь с альбумином (Alb / альбумин) и остается связанным с Alb до захвата гепатоцитами. Bu относительно нерастворим в воде до связывания с Alb.
- Когда Bu попадает в печень и ее проницаемые для белков синусоиды, он, вероятно, связывается с мембранными рецепторами гепатоцитов семейства органических анион-транспортирующих полипептидов (OATP, Organic Anion Transporting Polypeptide), проникает в гепатоциты без Alb и связывается с Y-белком (лигандином), глутатион-трансферазой (GST, Glutathione S-Transferase) или с Z-белком (белком, связывающим жирные кислоты). Этот процесс проникновения Bu в гепатоциты является пассивным, но облегченным; связывающие белки усиливают процесс, уменьшая обратный отток Bu обратно в синусоидальную плазму.
- Внутри гепатоцитов Bu диффундирует к эндоплазматическому ретикулуму, где он конъюгируется с глюкуронидом (или более чем на 60% с глюкозой у лошадей) с образованием моноглюкуронида билирубина или диглюкуронида билирубина, в совокупности называемых Bc (Bilirubin conjugated / билирубин конъюгированный). Конъюгация делает молекулу более растворимой в воде, и она диффундирует к канальцевой мембране.
- Bc транспортируется из гепатоцитов в канальцы (лимитирующая скорость стадия экскреции билирубина) с помощью энергозависимой транспортной системы для органических анионов, отличных от желчных кислот (BAs). Основным транспортером, вероятно, является белок 2, ассоциированный с множественной лекарственной устойчивостью (MRP2, Multidrug Resistance-associated Protein 2), также называемый канальцевым мультиспецифичным транспортером органических анионов (cMOAT, canalicular Multispecific Organic Anion Transporter). Имеются данные, что часть Bc перемещается из гепатоцитов в синусоидальную кровь (через MRP3, Multidrug Resistance-associated Protein 3) и обратно в гепатоциты через OATP перед секрецией.
- Bc в желчи попадает в кишечник и расщепляется до уробилиногена (Ub), который бесцветен. Уробилиноген может пассивно всасываться в кишечнике, а затем попадать в гепатоциты для экскреции с желчью или миновать печень и выводиться с мочой. Уробилиноген также может расщепляться до стеркобилиногена (Sb), который имеет темно-коричневый цвет и выводится с калом.
- Bc, который переходит из гепатоцитов в кровь без повторного захвата, может пройти через клубочковый фильтрационный барьер и выводиться с мочой. Поскольку альбумин (Alb) не проходит через клубочковый фильтрационный барьер у большинства млекопитающих, комплекс Bu/Alb не попадает в мочу у этих животных.
Bu/Alb — билирубин, связанный с альбумином; UDP-G (Uridine Diphosphoglucuronide / уридиндифосфоглюкуронид) — уридиндифосфоглюкуронид.

Рисунок 3.4. Кинетика Fe у здоровых животных.
• Усвоение: Рацион домашних млекопитающих может содержать Fe2+ или Fe3+. Поступивший в организм Fe3+ преобразуется в Fe2+
с помощью фермента редуктазы железа (цитохрома в двенадцатиперстной кишки или Dcyt b), поверхностного фермента, перед поступлением в энтероциты через
транспортер 1 двухвалентного металла (DMT1). Ферропортин и гефестин переносят Fe2+ в трансферрин плазмы в форме Fe3+.
• Транспорт: Почти все железо в плазме крови связывается с апотрансферрином, транспортным белком (β-глобулином), вырабатываемым гепатоцитами. Когда железо связывается с апотрансферрином, этот комплекс называется трансферрином. Трансферрин переносит Fe3+ в ткани и из них для использования клетками или для хранения. В здоровом организме около трети мест связывания железа с трансферрином занято железом. Многие клетки имеют рецепторы к трансферрину; важно отметить, что к ним относятся предшественники эритроидов.
• Использование в эритроидных предшественниках: После того, как трансферрин связывается с эритроидными предшественниками и попадает в них, Fe3+ диссоциирует из апотрансферрина в ферритин или включается в гем (Fe2+), а затем в гемоглобин. Большая часть апотрансферрина не подвергается деградации и возвращается в плазму. Предшественники эритроидов могут также получать железо из макрофагов-кормилиц, которые используют несколько механизмов получения железа, в том числе с помощью трансферрина.
• Хранение: Fe3+ содержится в двух белково-железистых комплексах: ферритине (плазма и ткани) и гемосидерине (тканевые макрофаги).
⚬ Ферритин состоит из апоферритина, образованного в комплексе с Fe3+, и является относительно растворимым и подвижным источником Fe3+. Существует несколько форм апоферритина из-за различных комбинаций H- или L-субъединиц. Ферритин плазмы - это гликозилированный полимер, который относительно беден железом. Тканевой ферритин, который является негликозилированным и относительно богатым железом, вырабатывается многими клетками, в первую очередь макрофагами, гепатоцитами, эпителиоцитами слизистой оболочки кишечника и предшественниками эритроидов. Синтез апоферритина гепатоцитами и макрофагами усиливается при воспалении и увеличении запасов железа. Апоферритин - это положительный белок острой фазы, который измеряется и регистрируется как
[ферритин].
⚬ Гемосидерин — относительно нерастворимый, малоподвижный источник Fe3+, представляющий собой основную форму хранения железа, количество которого увеличивается при избытке железа. Гемосидерин — это комплекс белка и оксидов железа, содержащийся в лизосомах макрофагов. Ферритин служит источником железа в гемосидерине, и железо, хранящееся в гемосидерине, может переноситься в ферритин для дальнейшего использования. Макрофаги, содержащие гемосидерин, присутствуют в селезенке, печени и костном мозге большинства млекопитающих, но костный мозг здоровой кошки не содержит гемосидерина, по крайней мере, в недостаточном количестве, чтобы его можно было обнаружить обычными методами окрашивания на железо.
• Тканевые формы: относительно небольшое количество железа присутствует в миоглобине, каталазе, пероксидазах и цитохромах, а также в ферритине и гемосидерине.
A. Общее количество железа в организме распределяется в трех основных местах, отвечающих за состояние здоровья: (i) около 50-70 % содержится в эритроцитах, (ii) около 25-40 % накапливается в организме (у молодых животных его мало) и (iii) остальная часть содержится в других молекулах (например, миоглобине, цитохромах и ферментах). Очень малая доля, которая циркулирует в плазме в виде трансферрина, не отражает общих запасов железа в организме. Fe-связывающие белки играют важную роль в ограничении свободного железа и связанных с ним окислительных повреждений, обеспечивая при этом биологическую доступность железа там, где оно необходимо.
B. Физиологические процессы или концепции представлены на рисунке 3.4.
C. У большинства млекопитающих скорость всасывания железа в кишечнике зависит от потребности организма в железе, т.е. при необходимости железа усваивается больше. У здоровых взрослых особей количество железа, поглощаемого за день, составляет очень небольшой процент от общего запаса железа в организме. Гораздо большее количество доступного железа образуется в результате продолжающейся деградации стареющих эритроцитов, каждый из которых содержит много миллионов атомов железа; это обеспечивает непрерывный приток переработанного железа, большая часть которого используется для синтеза Hgb в эритроидных предшественниках.
D. Всасывание железа в кишечнике и его высвобождение из клеток регулируются гепсидином, небольшим пептидным гормоном, который синтезируется преимущественно гепатоцитами. Гепсидин собак имеет молекулярную структуру, сходную с гепсидином человека.
1. При низких концентрациях гепсидина в энтероцитах усиливается синтез мембранного транспортера железа ферропортина, который в сочетании с гефестином, медьсодержащей железоксидазой, связанной с церулоплазмином, транспортирует Fe2+ в трансферрин плазмы в форме Fe3+.
2. Напротив, повышенный уровень внутриклеточного [гепсидина] снижает уровень функционального мембранного ферропортина, а также вызывает деградацию апикального транспортера железа. Следовательно, эпителиальные клетки кишечника поглощают меньше железа и меньше его может перемещаться из энтероцитов в плазму; неусвоенное железо, содержащееся в энтероцитах, теряется при отслаивании и замене клеток слизистой оболочки.
3. Действие гепсидина на макрофаги аналогично блокированию и усвоению ферропортина, так что экспорт железа снижается, например, в nRBCs (ядерные эритроциты) для синтеза Hgb.
E. Синтез гепсидина усиливается при повышении доступности железа или увеличении выработки IL-6
во время воспаления, и он косвенно снижается за счет увеличения [Epo].
1. Увеличение запасов железа в тканях и увеличение содержания железа в плазме (связанного с трансферрином железа) ощущается печенью и приводит к усилению выработки гепатоцитами гепсидина, что снижает всасывание железа, когда в нем нет необходимости. Дефицит железа снижает синтез гепсидина, тем самым увеличивая всасывание железа, когда это необходимо.
2. Анемия или другие причины почечной гипоксии вызывают повышение содержания эритропоэтина в плазме крови. Эпо стимулирует предшественников эритроидов вырабатывать эритроферрон, гормон, который подавляет выработку гепатоцитами гепсидина и, таким образом, делает железо более доступным для эритропоэза. При усилении эритропоэза количество циркулирующего [железа] уменьшается, что также снижает передачу сигналов для выработки гепсидина. В результате увеличивается поступление железа из энтероцитов в плазму и увеличивается высвобождение железа из макрофагов в плазму и предшественники эритроцитарной массы.